
Scientific update

Eco-evolutionary dynamics of living systems: Theory

Many real-world biological mechanisms can be properly represented mathematically only in terms of measure-valued processes. Drawing on recent mathematical developments, Evolution and Ecology Program (EEP) researchers have underpinned and extended classical methods to deal with such more realistic ecological situations.
In particular, scientists respectively underpinned models of the deterministic theory of physiologically structured populations and the adaptive dynamics describing clonal and one-locus Mendelian evolution [1] [2] [3].
Finally, after 25 years of successful use, it was proven that the so-called escalator boxcar train method for numerically solving physiologically structured population models has indeed the required convergence property [4] .
A number of efforts have extended the reach of the canonical equation of adaptive dynamics theory to function-valued traits [5], to complex interaction structures [6], to physiologically structured populations with multi-locus Mendelian genetics [7], and to research on stochastically fluctuating environments [8].
Gratifyingly, in all these cases the extensions have kept the basic form of the equation intact, with only some terms having to be modified with scalar factors to account for the additional complexity.
In view of the current unprecedented expansion of model-based analyses, it is now imperative to derive high-level rules that capture the essentials of large classes of models in one go.
In this spirit, it was shown how a concept of individual focusing on relations used in modeling instead of on ontological perceptions, can aid in spotting simple mathematical relationships in complicated ecological and evolutionary models [9].
Scientists derived the necessary and sufficient conditions under which life-cycle graphs allow simple interpretable expressions for the average number of offspring an individual produces over its lifetime [10]. Such expressions were used to elucidate what the structure of an organism’s life-cycle graph can already tell its life-history evolution [11].
To better understand limitations on the adaptability of ecological traits, researchers examined how mechanical constraints affect spine evolution [12]. Finally, it was shown how size-dependent energy acquisition, biomass production, and mortality rates, lie at the heart of various population dynamical and community dynamical phenomena [13].
One of the key features of adaptive dynamics theory is that it is the only theoretical framework that on its own, i.e., without invoking additional ad hoc hypotheses, predicts the generation of biological diversity.
Simple rules for predicting such generation in evolutionary models allowing for finite mutational steps were developed [14]. It was also elucidated how patterns of diversity arising under competitive interactions depend on the shape of the underlying competition functions [15] (Figure 1).
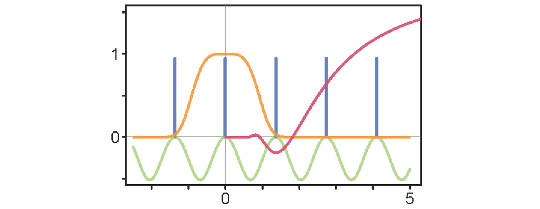
Figure 1. Example of an innovative mathematical analysis elucidating biodiversity patterns. In ecology, competition functions (orange) describe how the intensity of competition among individuals changes with their spatial or phenotypic distance (along the horizontal axis). Understanding how the Fourier transform (red) of such a function affects the resultant fitness landscape (green) enables predicting that clustered populations and/or multiple species (blue) arise if the Fourier transform has a negative minimum, with the wavelength at this minimum determining the inter-cluster gap.
A study eased the access to adaptive dynamics theory for a larger public by summarizing its essential features and reviewing its relationship with related frameworks, illustrated through didactically chosen examples [16].
References
[1] Metz JAJ & Tran VC (2013). Daphnias: From the individual based model to the large population equation. Journal of Mathematical Biology 66: 915–933.
[2] Gupta A, Metz JAJ & Tran VC (2013). A new proof for the convergence of an individual based model to the trait substitution sequence. Acta Applicandae Mathematicae, in press. doi: 10.1007/s10440-013-9847-y.
[3] Collet P, Méléard S & Metz JAJ (2013). A rigorous model study of the adaptive dynamics of Mendelian diploids. Journal of Mathematical Biology 67: 569–607.
[4] Brännström Å, Carlsson L & Simpson D (2013). On the convergence of the Escalator Boxcar Train. SIAM Journal of Numerical Analysis 51: 3213–3231.
[5] Parvinen K, Heino M & Dieckmann U (2013). Function-valued adaptive dynamics and optimal control theory. Journal of Mathematical Biology 67: 509–533.
[6] Allen B, Nowak MA & Dieckmann U (2013). Adaptive dynamics with interaction structure. American Naturalist 181: E139–E163.
[7] Metz JAJ & de Kovel CGF (2013). The canonical equation of adaptive dynamics for Mendelian diploids and haplo-diploids. Interface Focus 3: 20130025.
[8] Ripa J & Dieckmann U (2013). Mutant invasions and adaptive dynamics in variable environments. Evolution 67: 1279–1290.
[9] Metz JAJ (2013). On the concept of individual in ecology and evolution. Journal of Mathematical Biology 66: 635–647.
[10] Rueffler C & Metz JAJ (2013). Necessary and sufficient conditions for R0 to be a sum of contributions of fertility loops. Journal of Mathematical Biology 66: 1099–1122.
[11] Rueffler C, Metz JAJ & Van Dooren TJM (2013). What life cycle graphs can tell about the evolution of life histories. Journal of Mathematical Biology 66: 225–279.
[12] Galis F, Carrier DR, van Alphen J, van der Mije SD, Metz JAJ & ten Broek CMA. Running for life: Developmental and biomechanical constraints on homeotic transformations, in revision.
[13] De Roos AM, Metz JAJ & Persson L (2013). Ontogenetic symmetry and asymmetry in energetics. Journal of Mathematical Biology 66: 889–914.
[14] Ito HC & Dieckmann U (2013). Evolutionary branching under slow directional evolution. Journal of Theoretical Biology, in press. doi: 10.1016/j.jtbi.2013.08 .028.
[15] Leimar O, Sasaki A, Doebeli M & Dieckmann U (2013). Limiting similarity, species packing and the shape of competition kernels. Journal of Theoretical Biology 339: 3–13.
[16] Brännström Å, Johansson J & von Festenberg N (2013). The hitchhiker’s guide to adaptive dynamics. Games 4: 304–328.

CONTACT DETAILS
Principal Research Scholar Exploratory Modeling of Human-natural Systems Research Group - Advancing Systems Analysis Program
Principal Research Scholar Systemic Risk and Resilience Research Group - Advancing Systems Analysis Program
Principal Research Scholar Cooperation and Transformative Governance Research Group - Advancing Systems Analysis Program
Research program